
1 引言
陆面过程是影响天气和气候变化的基本物理、 生化过程之一, 它既包括生物物理过程(陆气之间的能量交换、 水汽交换和动量交换), 也包括生物化学过程(陆气之间的碳、 氮、 磷、 硫等物质循环)(陈海山等, 2002; 孙菽芬, 2005)。在众多地表类型中, 植被占陆地表面积的一半左右, 它通过影响陆面与大气间物质和能量的交换影响着天气和气候变化(戴永久等, 1996; 朱晗晖等, 2018)。因此, 深入研究陆面过程中的植被结构与功能, 是提高气候模式模拟精度及开展气候变化研究的重要内容(Betts, 1997; 牛国跃等, 1997)。其中, 根系作为植被的重要组成部分, 例如森林植被类型中根系占植被总鲜重20%~38%(Mokany et al, 2005), 根系在各种生态系统中发挥着重要的作用, 包括反映植被生长状况、 水分吸收和养分汲取等(Jackson et al, 1997; Quijano, 2012)。根系是土壤-植被-大气系统的重要纽带, 根系参数也是提高陆面模式乃至气候模式模拟能力的关键参数(Kleidon et al, 1998; Laio et al, 2006; Schneider et al, 2010)。
植被地上部分结构及功能长期以来受到广泛关注, 而地下根系部分却较少受重视(Copley, 1996; 黄建辉等, 1996; 王玉阳等, 2017)。根系深度和根系分布是用来反映根系状况及其功能的差异性的两个主要参数, 现有陆面模式对根系的描述大多都是围绕这两方面展开的(Arora et al, 2003; 吉喜斌等, 2006)。陆面模式BATS(Biosphere-Atmosphere Transfer Scheme)(Dickinson et al, 1993), TEM 4(Terrestrial Ecosystem Model)(McGuire et al, 1997), NOAH-LSM(Community Noah Land Surface Model)(Mitchell et al, 2005)等采用均一化根系分布方案, 每层土壤中的根系分布只与土壤层厚度与总深度有关: , 其中: 表示每层土壤中根系占总根系的比例, 用来表征根系分布; 表示每层土壤厚度; 表示土壤总深度。Gale et al(1987)提出一种静态渐近线式的根系分布方案: , 为深度z以上根系占总根系的比例, 即累积根系比例, 表征根系分布; 是与植被类型有关的衰减系数。这种根系分布方案被大多数陆面模式采用, 如NCAR-LSM(NCAR Land Surface Model)(Bonan, 1996), SiB(Simple Biosphere Model)(Sellers et al, 1986)。Zeng et al(2001)建立了一种呈指数形式衰减的根系垂直分布方案, 并采用依赖于植被类型的形态参数, 即: , 表示在深度为z处的累积根系比例 , a和b为形态参数并由查表方式获得, 这种方案在CLM4.0(Community Land Model)(Oleson et al, 2010), CLM4.5(Oleson et al, 2013)中得到应用。SSiB4(Simplified Simple Biosphere Model)(Xue et al, 1991)模式仅考虑土壤层厚度与根系总深度有关, 不同植被类型对应不同的土壤分层结构, 并给定固定不变的根系深度, 以此得到根系分布比例: , 其中: 为每层土壤中根系长度(深度); 为根系分布比例。以上方案虽然对根系分布作出不同形式的拟合, 但相比地上部分以叶面积指数、 茎高度指数等描述叶和茎的变化, 在制约因素上考虑不够充分, 不考虑外部环境对根系的影响, 导致这些参数化方案得到的根系分布是静态的, 不能反映植被对环境的适应性(才奎冶等, 2015)。
根系是植被获取水分及养分的重要器官, 根系分布也决定着植被从不同土壤层中吸取水分及养分的能力(李凯等, 2015; 张经天等, 2019), 因此可以用土壤水分和养分状况来表征根系分布状况(郭京衡等, 2016)。土壤水分不足或过量时对根系生长和叶片气孔导度产生的限制称为土壤水分胁迫, 土壤养分不足或过量时对根系生长的限制称为土壤养分胁迫, 本研究中土壤养分由土壤氮含量进行表征。此外, 陆面模式还采用净增长生物量在各个组织器官间的分配描述植物生长, 因此用每层土壤中根系碳量的比例表征根系分布。本研究考虑根系碳量、 土壤水分胁迫及土壤养分(氮)胁迫, 在陆面模式SSiB4中构建动态根系方案, 研究根系动态过程对改进陆面模式模拟效果的影响, 并探讨其影响机制, 为提高陆面模式的模拟能力提供参考。
2 模式、 方案介绍及数据来源
2.1 SSiB4/TRIFFID介绍
SSiB4/TRIFFID(Top-down Representation of Interactive Foliage Including Dynamics, TRIFFID)是包含了动态植被模块并完全耦合地表能量循环、 水循环和碳循环的陆面模式, 其模拟表现已经通过在不同气候和土地覆盖条件下的多点测评(Cox, 2001; Zhan et al, 2003; Zhang et al, 2015)。在与动态植被模块耦合时, SSiB4模拟植物净光合作用同化率、 自养呼吸和其它地表条件(如植被冠层温度、 气孔导度等), 并将土壤温度、 土壤湿度及植物净光合同化量传递给TRIFFID, TRIFFID将模拟的同化量分配给模拟区域内存在的各种植被类型, 通过计算植被参数(如植物高度和LAI), 来模拟各种植被的生长情况, 还包含动态竞争机制, 以实现不同植被类型之间的生长和竞争。最新版本的SSiB4/TRIFFID中, 将全球植被分为7大类: 常绿阔叶林、 北方阔叶林、 针叶林、 C3植物、 C4植物、 苔原和灌木, 此外还包括裸地。
图1给出了SSiB4/TRIFFID计算流程, TRIFFID将每种植被总碳量, 按照异速生长关系分配到叶、 茎、 根三个器官, 即得到根系总碳量。SSiB4/TRIFFID能够表征的碳、 水相关的参数, 是进行碳氮水相互作用研究及动态根系方案制定的基础。关于对根系的土壤养分胁迫, 则由外部读入的土壤全氮数据实现。
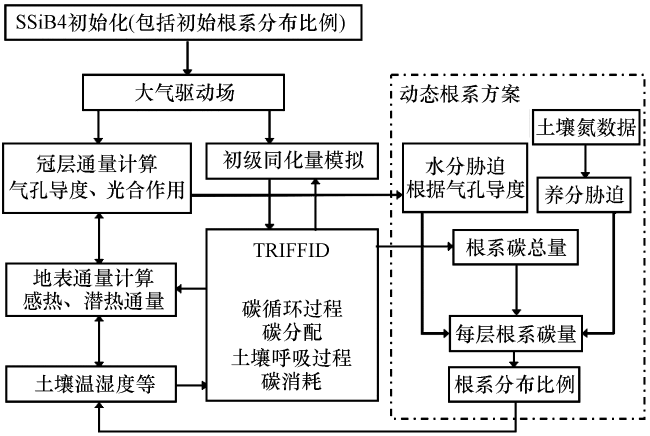
2.2 考虑碳氮水过程的动态根系方案
原版本的SSiB4/TRIFFID模式采用均一化根系分布方案, 根系分布不随外界条件变化, 不能反映植被对环境的适应性, 考虑到植被根系获取水分及养分的功能, 本文参考一种考虑土壤水分胁迫及氮胁迫的碳分配方案(Hatzis, 2010; 王媛媛, 2016), 每层土壤中根系碳分配量为:
,
式中: 为第i层的土壤厚度; 为第i层土壤中的植被萎蔫系数; 为第i层土壤中的氮含量相对比例(即土壤养分胁迫系数); 为该土壤柱的水分胁迫系数; 为根系总碳量; 为第i层土壤中根系碳量。
SSiB4/TRIFFID模式为三层土壤结构, 根系仅分布在第二(浅层)层和第三层(深层), 且不存在土壤柱的水分胁迫系数 的模拟, 为适应模式将式(1)修改为以下形式:
,
,
,
式中: 为第i层土壤中的氮水胁迫系数; r为该土壤柱中的氮水胁迫系数。
,
式中: 和 分别是叶片气孔完全关闭和打开时的土壤水势(单位: mm); 是第i层土壤基质势(单位: mm), 、 是经验系数。
由于TRIFFID直接模拟植被根系碳量, 因此设置为期十天的循环周期, 模拟根系自然生长状态, 将根系碳量分配与根系分布比例联系在一起:
,
,
计算出根系分布比例为:
,
式中: 为n时刻第i层土壤中的根系分布比例。
在运行模式前首先进行模式变量初始化, 包括对各个植被类型的土壤每层根系分布比例的初始化, 读入的是CLM4.0模型采用的指数衰减方案中的对应常数。每个网格上, SSiB4对不同类型植被进行计算, 采用自上而下(从植被到土壤)的顺序展开。首先计算冠层通量(如气孔对水汽及二氧化碳的阻抗、 感热通量和光合作用等), 然后计算土壤中各种物理过程, 得到各层土壤湿度及萎蔫系数。基于SSiB4计算得到的不同植被类型的地表参数和土壤参数, TRIFFID依照异速生长关系计算出根系碳量, 然后从SSiB4中读入土壤水分胁迫及根系碳量, 并从外部文件读入土壤氮胁迫系数, 按公式(6) ,(7) 进行每层土壤中根系碳量分配, 接着对土壤柱水分胁迫系数、 累积碳量计算及根系分布比例进行更新, 作为根系动态变化的表征返回给SSiB4, 用于根系吸水的计算及土壤湿度的更新。
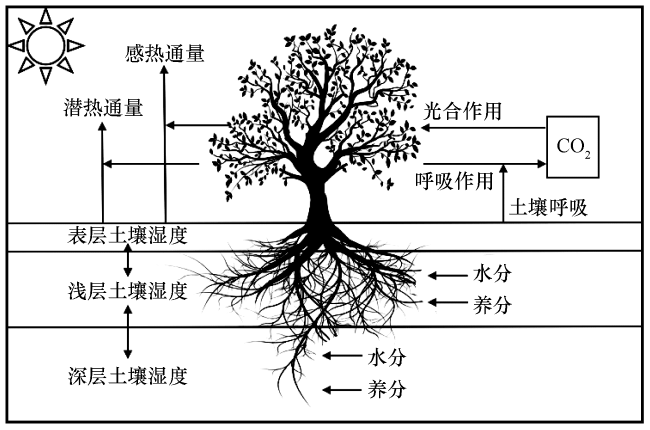
考虑动态根系变化后, 将对模式中的物质循环过程和水热循环过程产生影响, 具体影响机制如图2所示。每层土壤中根系分布比例的动态变化, 首先导致根系在每层土壤中吸水量的变化, 然后引起每层土壤湿度的变化。土壤湿度作为最重要的地表变量之一, 会对陆地生态系统中的物理和生化过程产生重要作用(马英赛等, 2019)。动态根系变化通过影响土壤湿度来调节植被生理生化过程(如蒸腾作用), 从而影响二氧化碳通量, 进而改变表层土壤蒸发、 植被反照率等地表参量, 改变地表净辐射在感热和潜热通量之间的分配(刘双等, 2018; 陈银萍等, 2019)。
2.3 资料来源
选取两个典型的设立在森林生态系统中的站点作为模式的验证站点, 这两个站点分别是位于亚马逊热带雨林中的BRSa3站点和位于中国江西省千烟洲的CN-Qia站点。除必须的驱动变量用来驱动离线陆面模式外, 这两个站点还提供了用以评估模式模拟能力的变量, 包括土壤湿度、 二氧化碳通量、 潜热通量和感热通量。
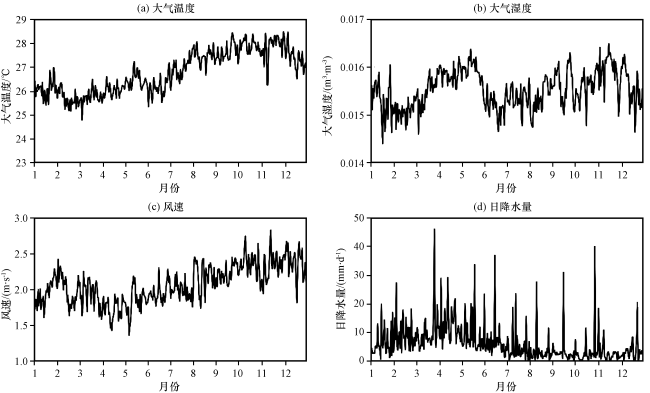
亚马逊地区拥有世界50%的热带雨林, 对全球氮水循环至关重要, 陆地生态系统的生理和生化过程会显著影响区域和全球气候, 因此选用这一地区进行单点模拟试验(Avissar et al, 2002)。BRSa3站点(http: //sites.fluxdata.org/BR-Sa3/)位于巴西塔帕若斯河国家森林公园(3.02°S, 54.97°W), 下垫面主要植被类型为热带常绿阔叶林。从2000 -2004 年BRSa3站点日平均大气温度、 比湿、 风速和降水的情况(图3)可以看出, 该站点多年平均2 m气温为26 ℃, 最高气温34 ℃, 最低气温20 ℃, 气温的季节变化较小。多年平均总降雨量为2044 mm, 7 -12月高温少雨为干旱季, 具有典型的热带雨林气候特征。
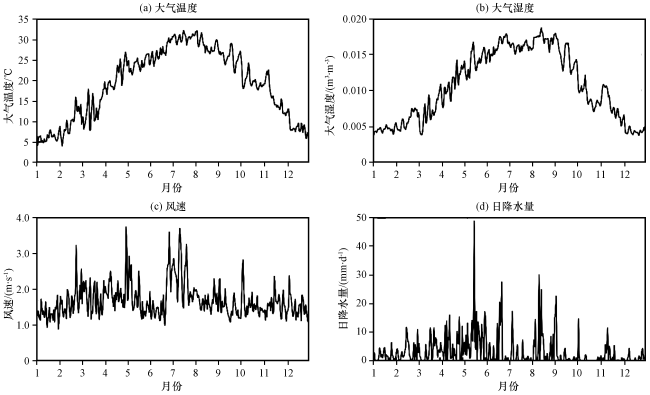
千烟洲地处南方丘陵区, 经过多年治理, 原本破坏殆尽的植被恢复为以人工林或草、 灌丛次生植被为主, 形成农田、 森林等典型生态系统结构, 可以更好地进行生态系统物质循环、 能量流动研究(刘允芬等, 2004)。CN-Qia站点(http: //sites.fluxdata.org/CN-Qia/)位于中国江西省千烟洲(115.03°E, 26.44°N), 下垫面主要植被类型为人工针叶林。从2003 -2005年CN-Qia站点日平均2 m温度、 比湿、 风速和降水情况(图4)可以看出, 该站点多年平均气温为18 ℃, 最高气温40 ℃, 最低气温-5 ℃, 多年平均总降雨量为1169 mm, 夏季高温多雨, 具有典型的亚热带季风气候特征。
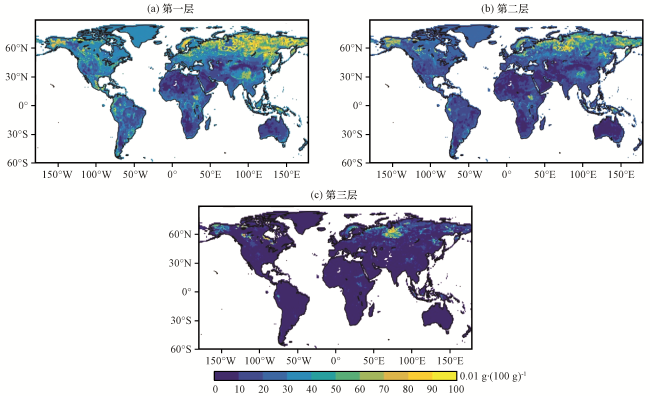
北京师范大学发布了一套网格化的土壤数据集(The Global Soil Dataset for Earth System Modeling, GSDE)(Shangguan et al, 2014), 此数据集基于世界土壤图和多个区域和国家级土壤数据库, 可以用于地球系统模型。本研究采用GSDE的土壤全氮含量[Total Nitrogen, 0.01 g·(100g)-1]数据为模式提供土壤氮含量, 该数据的空间分辨率为30弧秒, 垂直分为8层。在SSiB4/TRIFFID中, 土壤垂直结构分为3层, 每一层的厚度依赖于初始化网格上的主导植被类型, 第一层厚度一般是0.02 m, 第二层在0.17~1.48 m间变化, 第三层在0.3~2 m间变化。为了使GSDE的土壤全氮含量与SSiB4/TRIFFID的土壤厚度保持一致, 将土壤全氮含量在垂直方向线性插值到模式的土壤层上, 并在水平方向上进行空间升尺度, 得到全球范围内空间分辨率为1°×1°的3层土壤氮含量数据[图5, 基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1667的标准地图制作, 底图无修改]。该数据作为外部文件写入SSiB4/TRIFFID中, 用来表征土壤氮胁迫系数, 供动态根系模块调用。
2.4 试验设计
模式积分步长为3 h, 所有驱动场数据都是从记录时刻开始的步长时间段内的平均值, 模拟结果每3 h输出一次, 参与验证的模拟结果都是直接输出值的平均值。驱动数据包括2 m气温(单位: K)、 气压(单位: Pa)、 大气湿度(单位: m³·m-3)、 10 m风速(单位: m·s-1)、 入射长短波辐射(单位: W·m-2)和降水(单位: mm·s-1), 用来驱动离线陆面模式。本研究设计了两组试验: (1)控制试验(CTL试验), 使用原SSiB4/TRIFFID模式, 根据站点真实情况调节土壤孔隙度、 初始根系分布比例、 初始植被覆盖率及主导植被类型。(2)对比试验(NEW试验), 使用加入动态根系方案的SSiB4模式, 其余设置与CTL试验相同。观测数据结果记为OBS。另外, SSiB4/TRIFFID模式在两站点的土壤垂直分层相同, 即表层为0~0.02 m, 浅层为0.02~1.5 m, 深层为1.5~3.5 m, 由于观测数据有限, 在对比观测和模拟的土壤湿度时使用0.2 m观测值与浅层土壤湿度模拟值。
3 结果分析
3.1 BRSa3站点结果分析
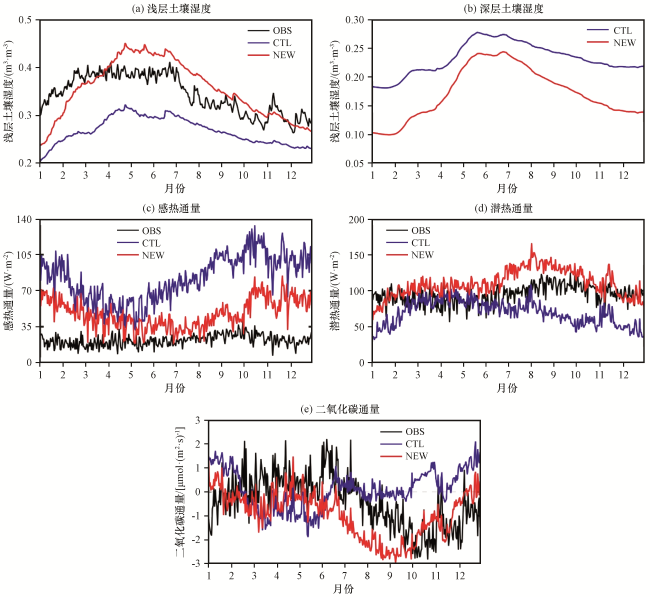
图6 2000 -2004年BRSa3站点各变量的日平均模拟值与观测值的比较Fig.6 Daily averaged variables of BRSa3 station compared with the observed from 2000 to 2004 |
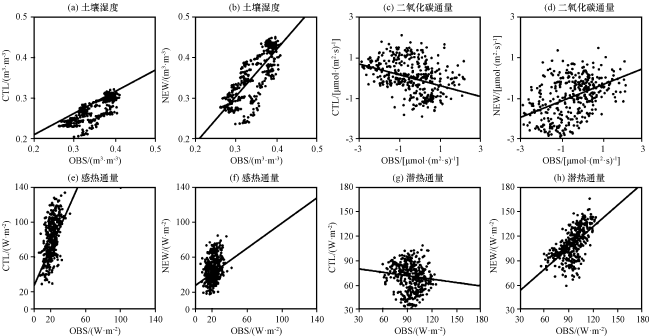
图7 2000 -2004年BRSa3站点模式结果与观测值散点分布比较Fig.7 Scatter plots of BRSa3 station compared with the observed from 2000 to 2004 |
表1 两站点各植被类型覆盖度与改进前后根系分布比例Table 1 Coverage rate and root distribution ratio of CTL and NEW simulation in two site stations |
| 站点 | 变量 | 常绿阔叶林 | 针叶林 | 阔叶林 | C3草地 | C4草地 | 灌木 | 苔原 | 裸地 |
|---|---|---|---|---|---|---|---|---|---|
| BR站 | 覆盖度 | 0.95 | - | - | - | - | - | - | 0.05 |
| CTL浅层根系浅层 | 1.00 | 0.98 | 0.94 | 1.00 | 1.00 | 1.00 | 0.17 | ||
| NEW浅层根系 | 0.65 | 0.76 | 0.81 | 0.82 | 0.91 | 1.00 | 0.11 | ||
| CTL深层根系 | 0.00 | 0.02 | 0.06 | 0.00 | 0.00 | 0.00 | 0.83 | ||
| NEW深层根系 | 0.35 | 0.24 | 0.19 | 0.18 | 0.09 | 0.00 | 0.89 | ||
| CN站 | 覆盖度 | 0.14 | 0.68 | - | 0.05 | 0.10 | - | - | 0.03 |
| CTL浅层根系浅层 | 1.00 | 0.98 | 0.94 | 1.00 | 1.00 | 1.00 | 0.17 | ||
| NEW浅层根系 | 0.55 | 0.42 | 0.91 | 0.85 | 0.88 | 1.00 | 0.29 | ||
| CTL深层根系 | 0.00 | 0.02 | 0.06 | 0.00 | 0.00 | 0.00 | 0.83 | ||
| NEW深层根系 | 0.45 | 0.58 | 0.09 | 0.15 | 0.12 | 0.00 | 0.71 |
— 表示该项数值小于0.01; 空白表示无根系分布 |
表2 两站点统计特征Table 2 Statistical characteristics of two site stations |
| 站点 | 变量 | 平均偏差 | 均方根误差 | 平均值 | 相关系数 | ||||
|---|---|---|---|---|---|---|---|---|---|
| CTL | NEW | CTL | NEW | CTL | NEW | CTL | NEW | ||
| BR站 | 浅层土壤湿度/(m³·m-³) | 0.08 | 0.03* | 0.08 | 0.04 | 0.27 | 0.36 | 0.76 | 0.78 |
| 二氧化碳通量/[µmol·(m2·s)-1] | 1.39 | 0.03* | 1.64 | 1.24 | 0.08 | -0.95 | -0.34 | 0.42 | |
| 感热通量/(W·m-2) | 60.23 | 25.78* | 63.42 | 29.21 | 81.94 | 47.44 | 0.54 | 0.25 | |
| 潜热通量/(W·m-2) | 27.85 | 19.26* | 33.74 | 21.87 | 69.20 | 110.1 | -0.14 | 0.61 | |
| CN站 | 浅层土壤湿度/(m³·m-³) | 0.034 | 0.029* | 0.039 | 0.034 | 0.23 | 0.27 | 0.71 | 0.75 |
| 二氧化碳通量/[µmol·(m2·s)-1] | 1.14 | 0.63* | 1.50 | 0.76 | -2.14 | -1.88 | 0.61 | 0.74 | |
| 感热通量/(W·m-2) | 28.06 | 22.68* | 34.88 | 29.29 | 44.14 | 37.10 | 0.61 | 0.57 | |
| 潜热通量/(W·m-2) | 21.22 | 13.35* | 25.47 | 17.46 | 69.48 | 61.03 | 0.93 | 0.95 | |
星号表示与CTL相比通过99%显著性检验; 标粗数值为相关系数, 没有单位 |
加入动态根系方案后, 根系向深层发展, 吸收更多深层水分供给植被蒸腾, 导致浅层土壤湿度有所增加, 深层土壤湿度有所减少。NEW试验模拟出的土壤湿度有较大改进, 平均值由0.27 m³·m-³增加到0.36 m³·m-³, 平均偏差由0.08 m³·m-³降低至0.03 m³·m-³, 相关系数由0.76增加到0.78(均通过显著性检验, P<0.01), 更接近观测值。这是因为亚马逊地区主要植被类型为热带常绿阔叶林(表1), 实际根系多分布于深层, 原模式方案中根系大多分布在浅层, 无法模拟根系深层吸水需求。动态根系分布使得更多根系向土壤深层发展, 浅层吸水量下降而深层吸水量增长, 因此土壤浅层湿度增加, 更接近真实情况。
对于感热通量及潜热通量, CTL试验模拟结果与OBS表现有较大差异, 感热通量量级表现不佳, 潜热通量在干旱季趋势表现不一致。加入动态根系方案后, 土壤湿度的模拟有较大改善, 影响到土壤反照率、 土壤温度等地标参量, 通过土壤物理过程使得感热及潜热通量产生变化。NEW试验中, 上述偏差有一定程度改善, 平均偏差由60.23 W·m-2和27.85 W·m-2降低到25.78 W·m-2和19.26 W·m-2, 均方根误差由63.42 W·m-2和33.74 W·m-2降低到29.21 W·m-2和21.87 W·m-2(均通过显著性检验, P<0.01), 模拟精度仍有待提升。
对于二氧化碳通量, OBS表现为整体碳吸收, 平均值为-0.41 µmol·(m2·s)-1, CTL试验模拟结果整体表现却为碳排放, 与观测结果不符。加入动态根系方案后, 二氧化碳通量平均值由0.08 µmol·(m2·s)-1降低到-0.95 µmol·(m2·s)-1(均通过显著性检验, P<0.01), 整体趋势修正为碳吸收, 平均偏差由1.39 µmol·(m2·s)-1降低至0.03 µmol·(m2·s)-1, 相关系数由-0.34升至0.42, 更接近观测值。尤其将7 -12月干旱季CTL中表现为很强的碳排放修正为碳吸收, 干旱季二氧化碳通量平均值由0.35 µmol·(m2·s)-1降低到-1.56 µmol·(m2·s)-1, 与OBS平均值-1.1 µmol·(m2·s)-1更接近, 干旱季对整体时段绝对偏差改进的贡献率达76%, 相关系数由-0.1升至0.07, 以观测值衡量的绝对偏差的改进幅度为33%。这是因为动态根系使深层根系相对根量增多, 即使在干旱季浅层土壤水分供给不足的情况下, 也可以从深层土壤吸取水分以满足正常需求, 植被对季节性干旱的响应得到明显改善。根系动态分布对二氧化碳通量的影响主要是通过植被生理生化过程产生的, 这种对环境状况的适应性表现在二氧化碳通量仍保持为碳吸收状态。
3.2 CN-Qia站点结果分析
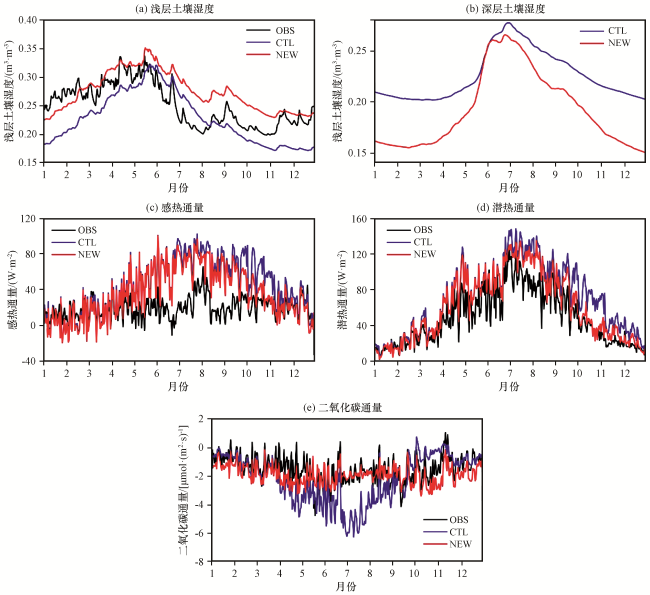
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
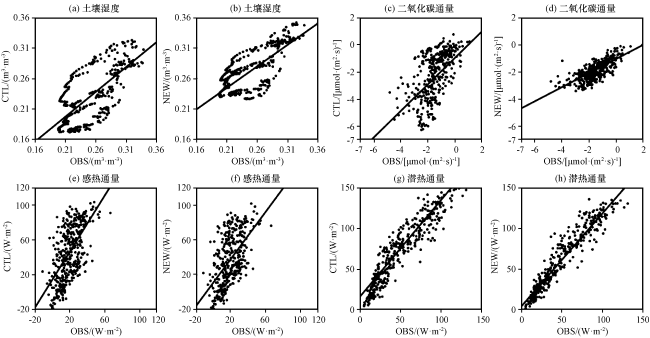
对于土壤湿度, CTL试验模拟结果与观测值有较好的一致性, 相关系数达0.71。加入动态根系方案后, NEW试验模拟出的浅层土壤湿度亦有改进, 相关系数提升至0.75, 平均值由0.23 m³·m-³增加到0.27 m³·m-³, 平均偏差由0.034 m³·m-³降低至0.029 m³·m-³(均通过显著性检验, P<0.01), 更接近观测值。CN-Qia站点的植被类型较为复杂(表1), 虽然主要植被类型为针叶林, 但是依然有草地、 灌木等浅根系植被分布。由于本研究没有对根系深度进行限制, 无法区分浅根系和深根系, 高估了对浅根系植被在深层土壤中的相对根量, 进而低估了浅层吸水量, 造成浅层土壤湿度模拟值部分偏大。
对于感热通量及潜热通量, 加入动态根系方案后, 由于对土壤湿度改善程度有限, 两者略有改善。平均偏差由28.06 和21.22 W·m-2降低到22.68和13.35 W·m-2, 均方根误差由34.88和25.47 W·m-2降低到29.29和17.46 W·m-2(均通过显著性检验, P<0.01)。
对于二氧化碳通量, OBS表现为整体碳吸收, 平均值为-1.47 µmol·(m2·s)-1, 加入动态根系方案后, NEW试验修正了对整体碳吸收能力的高估, 二氧化碳通量平均值由-2.14 µmol·(m2·s)-1升高至-1.88 µmol·(m2·s)-1, 平均偏差由1.14 µmol·(m2·s)-1降低至0.63 µmol·(m2·s)-1, 相关系数由0.61升至0.74(均通过显著性检验, P<0.01), 更接近观测值。特别在6 -8月夏季, NEW试验较好修正了模式对碳吸收能力的高估, 夏季二氧化碳通量平均值由-4.05 µmol·(m2·s)-1升至-1.97 µmol·(m2·s)-1, 更接近OBS平均值-1.93 µmol·(m2·s)-1, 夏季对整体时段绝对偏差改进的贡献率达44%, 以观测值衡量的绝对偏差的改进幅度为106%, 对二氧化碳通量量值修正明显。夏季虽然是植被生长旺盛的季节, 但不意味着不受环境及植被自身能力的制约, 如蒸腾作用要受到根系吸水量的限制。CTL试验高估了夏季根系供给蒸腾作用的根系吸水量, 表现在浅层土壤湿度在夏季迅速下降, 蒸腾作用强度被大大高估, 因此二氧化碳通量在夏季表现为强碳吸收, 与观测值差异较大。NEW试验修正了浅层根系吸水量, 使植被蒸腾量不至过大, 限制了夏季植被生长过分旺盛, 植被通过调节蒸腾作用适应环境变化, 大幅度提高二氧化碳通量, 表现为与观测结果一直的弱碳吸收状态, 更接近观测值。
4 结论与讨论
在陆面模式SSiB4/TRIFFID的基础上, 考虑了土壤水分胁迫、 土壤氮胁迫以及根系动态碳分配机制, 通过对位于亚马逊地区的BRSa3站点及中国江西千烟洲的CN-Qia站点进行单点模拟试验, 探讨根系动态变化对各关键变量模拟结果的影响, 得到以下结论:
(1) 浅层土壤湿度模拟精度提高, 因为更多根系生物量分配到深层, 根系由浅层向深层生长, 可以吸收更多深层水分用以植被蒸腾, 致使浅层土壤湿度有所增加且更接近观测值。两站点模拟精度提升程度有所差别, 原因可能在于两站点植被分布类型的不同。
(2) 二氧化碳通量模拟精度提高最为明显, 原因在于土壤湿度的变化最先影响植被生理生化过程, 直接作用于生态系统二氧化碳通量。两站点改善程度有所不同: 对于BRSa3站点, 将干旱季碳排放状态修正为碳吸收状态, 绝对偏差的改进幅度达33%; 对于CN-Qia站点, 将夏季强碳吸收状态修正为弱碳吸收状态, 绝对偏差的改进幅度达106%。这是因为植被在不同时期对环境状况的适应性在二氧化碳通量上的差异化体现。
(3) 潜热和感热通量与二氧化碳通量相比改善程度有限, 因为根区土壤湿度的变化最先通过与植被蒸腾作用相关的水分输运过程影响到植被生长, 其次还在浅层与表层土壤水势差的驱动下改变表层土壤湿度, 进而影响地表土壤蒸发等土壤物理过程, 影响其他地表通量。根区土壤湿度对地表潜热及感热通量的间接影响方式可能是造成上述差别的原因之一。
本研究仅在两个站点进行单点模拟试验, 未来需要搜集更多通量及根系观测资料, 进行更多验证分析, 甚至扩展到区域及全球试验; 仅进行离线模拟, 还应与地球系统模式耦合, 对植被根系动态变化的气候效应进行深入研究; 影响根系生长发育的因素还有很多, 本研究仅考虑碳氮水三方面的胁迫, 另外还有根系深度、 土壤盐分胁迫、 根系的水分再分配作用等等(Homaee et al, 2002; 阳园燕等, 2004), 需要完善其他影响因素对根系分布的调制作用。