

1 引言
植物的气孔调节是陆地生态系统碳-水耦合过程最重要的纽带(Buckley et al, 1999; Sperry et al, 2017), 充分理解气孔导度对环境因素的响应机制, 是植被和地球系统模型有效模拟生态系统碳、 水和能量平衡的必要条件(Jones, 1998; 罗丹丹等, 2019)。对于荒漠植物来说, 气孔调节是植物适应干旱逆境和调控蒸腾速率的主要机制(Schulze, 1986; 龚吉蕊等, 2005; 苏培玺, 2019; 罗欢等, 2020), 荒漠植物一方面通过自身的低水势和强大的根系从外界环境吸收水分, 另一方面又能在干旱环境中通过气孔运动机制来减少体内水分散失尽可能获取CO2(Chaves et al, 2002)。以往关于气孔导度对环境驱动响应的研究多集中在湿润环境中的树木, 对干旱生态系统特别是中亚地区的灌木研究较少(Zheng and Wang, 2014; Gu et al, 2018), 且国内外学者对于荒漠植物的研究主要涉及对光合和蒸腾作用的试验观测及对其变化过程和生理学意义的分析(苏培玺, 2019), 对于气孔导度的深入研究较少(袁国富等, 2012)。气孔导度变化极其复杂, 受到各环境因素的共同制约, 清楚了解气孔对环境驱动因子的响应有助于提升我们对蒸腾过程及其调控机制的认识。
陆地生态系统的水分有效性是决定植被生长和分布的重要因素之一, 特别是在干旱地区, 植物活动更是与之密切相关(苏培玺等, 2003; 王宇轩等, 2021)。降水和地下水是干旱区主要的水分来源, 目前关于降水对荒漠植物气孔导度的影响存在不同的看法, 一些研究认为降雨能明显改善荒漠水分环境, 影响植物生理生态过程和生态系统过程。如许皓和李彦(2005)的研究显示荒漠植物对降雨脉冲的响应不同, 能够利用较深层土壤水分的植物更具竞争优势, 其气孔导度对水分状况的敏感度较低, 利用浅层土壤水的植物对降雨的依赖性更强; Hoshika et al(2017)发现年平均降雨量会影响木本植物的最大气孔导度; Yu et al(2018)认为干旱胁迫下, 植物气孔导度随土壤水分的变化表现出明显规律, 降水变化将直接影响该地区的水分供应状态。也有研究表明沙漠灌木通常表现出非常深的根, 使它们能在深层土壤中汲取水分(Canadell et al, 1996), 对降雨引起的浅层土壤水分变化的生理响应并没有预期明显。如王亚婷和唐立松(2009)研究表明沙漠灌木体内的水分状况和生理状态对5 mm以下的降雨无显著响应; 曾晓玲等(2012)认为西北干旱区的天然降水无法满足植物的正常生长需求, 沙漠地下水位的变化在植被生长过程中发挥着重要作用。综上, 荒漠植物生理生态过程对外界环境水分变化的响应关系, 与植物根系分布和水分利用策略以及荒漠环境的地下水位和降雨大小等有关系, 降雨前后荒漠植物气孔导度的变化值得进一步研究, 而且目前关于气孔导度对综合环境因子响应研究较少, 限制了蒸腾模拟的能力, 所以有必要明确特定条件下植物种类对其周围环境因素的响应, 以解释外界环境因子和植物蒸腾速率以及气孔导度之间的关系, 理解干旱区植物的水分适应机制。
干旱区占地球陆地面积超过40%(Schimel, 2010), 干旱生态系统对区域或全球的能水循环及平衡有着重要影响。气孔导度是研究荒漠植被与大气间物质和能量交换的关键, 由于植物功能与大气和土壤的这种相互关系, 植物对水汽损失的控制也是生物圈和全球环流模式的重要组成部分(Berry et al, 2010; 高冠龙等, 2017; 李元寿等, 2019)。本文以河西走廊中部荒漠-绿洲过渡带C4植物梭梭(Haloxylon ammodendron)、 沙拐枣(Calligonum mongolicum)和C3植物泡泡刺(Nitraria sphaerocarpa)为研究对象, 主要探讨荒漠灌木叶片气孔导度对环境因子和叶片水势变化的响应规律, 及其对蒸腾速率的调控机理, 旨在深入理解荒漠植物对环境的适应机制, 为保护荒漠生态系统的稳定性以及荒漠-绿洲过渡带的植被恢复提供理论依据, 对干旱区的生态和环境建设具有基础的科学意义。
2 资料来源和方法介绍
2.1 试验区概况
试验区在中国科学院临泽内陆河流域研究站(39°20′50″ N, 100°07′48″ E; 海拔1384 m)绿洲-荒漠过渡带生态水文过程观测试验场, 位于河西走廊中部临泽绿洲北部, 毗邻巴丹吉林沙漠南缘。属于温带干旱荒漠气候类型, 年平均降水量约123 mm, 且主要集中于6 -9月, 年平均气温约8.9 ℃, 地下水位在4~4.5 m波动。植物生长季的总降水量为112.3 mm, 约占全年降水量的85.6%, 大部分单次降水量不超过6 mm, 平均值为3.7 mm。该过渡带主要以C4植物梭梭、 沙拐枣和C3植物泡泡刺3种荒漠灌木为优势种, 伴生有其他灌木[如: 红砂(Reaumuria soongorica)]、 半灌木[如: 合头草(Sympegma regelii)]、 草本[如: 盐生草(Halogeton glomeratus)]等。
2.2 观测方法
观测试验于2019 -2020年6 -9月展开, 3种荒漠植物的叶片气孔导度(gs, 单位: mol·m⁻2·s⁻1)、 蒸腾速率(Tr, 单位: mmol·m⁻2·s⁻1)、 光合有效辐射(PAR, 单位: µmol·m⁻2·s⁻1)、 饱和水汽压差(VPD, 单位: kPa)、 相对湿度(hs, 单位: %)和气温(T, 单位: ℃)均采用气体交换LI-6400XT便携式光合作用测定系统(LI-COR, Lincoln, NE, USA)的簇状叶室测量, 选择开放气路、 自然光源, 叶室上部装有光强传感器, 内部装有湿度和温度探头。测量时间段为07:00(北京时, 下同) -19:00, 每隔2 h测量一次, 根据当地天气预报, 在降雨前观测一天, 降雨后连续观测2天。每种植物随机选3个重复, 测量时用胶布将选取的同化枝固定并做好标记, 确保每次测量的叶面积相同, 测量结束后剪下叶片用扫描仪测量植物的叶面积, 由于扫描仪得到的为植物的投影面积, 而梭梭和沙拐枣的同化枝为圆柱体, 所以将得到的叶面积乘以π/2进行换算, 根据算出的实际叶面积重新计算叶片气孔导度和蒸腾速率。
植物叶片水势(Ψ, MPa)采用Model 1505植物压力室(PMS, Instrument Co, Albany, USA)测定, 日出前(06:00)测量凌晨水势, 之后与叶片气孔导度同步每隔2 h测量一次, 在选取的测量叶片气孔导度的植株上选距离地面高度相似、 长势相近的枝条(10 cm左右), 每次测3个重复。3种荒漠植物根区的土壤含水量(θ, 单位: m3·m⁻3)采用烘干法测定, 测量的土壤深度范围为0~200 cm, 每隔20 cm取一层土样。降水数据来自试验地小气候观测系统的降雨量传感器(TE525MM, Texas Electronics, Texas, USA), 位于植被冠层顶部。
2.3 灌木叶片气孔导度估算
气孔导度估算通常根据影响气孔行为的几个关键因素进行经验建模, 目前常用的气孔导度模型是Jarvis模型, 为多约束函数模型, 已被纳入到全球和区域的植被-气候模型(Hoshika et al, 2017)。Jarvis多变量响应函数乘积模型综合考虑了各因素(PAR、 VPD、 T和Ψ)对气孔导度的影响, 模型表达式为(Jarvis, 1976):
式中: gmax为生长季测定的环境条件最佳即光照充足、 饱和水汽压差较小、 温度适宜和植物根系层土壤较湿润时的最大叶片气孔导度值(Kelliher et al, 1995), 对于荒漠灌木最大叶片气孔导度的测定选取降雨后晴天的07:00 -10:00, 此时环境较为适宜, 短期气象条件影响较小, 本文梭梭、 泡泡刺和沙拐枣分别为0.237, 0.468和0.397 mol·m⁻2·s⁻1。f(PAR)、 f(VPD)、 f(T)和f(Ψ)分别表示气孔导度对光合有效辐射、 饱和水汽压差、 气温和叶片水势的响应函数, 数值为0~1。一些研究用Ψ表示土壤水分状态参数, 而已有研究表明这3种荒漠植物可以利用深层土壤水和地下水(Ji et al, 2016; Xu et al, 2017; 周海等, 2017), 深层土壤水的日变化较小, 对于日尺度的气孔导度模型来说, 并不能模拟出气孔导度响应土壤水分的日变化。气孔运动除了受到外界环境驱动外还与植物体内的水分平衡有关, 叶片水势(Ψ)能很好地反映植物的水分状况变化和土壤水分胁迫, 是更加直接有效和精确的参数(于贵瑞等, 2010)。利用2020年3种荒漠植物生长季内叶片气孔导度的实测数据, 用Matlab软件采用非线性最小二乘法拟合模型参数, 用2019年的数据验证模型。
3 结果与分析
3.1 降水对荒漠植物水分传输因子的影响
在自然状况下分析降水带来的影响, 降雨前后各指标的比较用双样本t检验, 各环境因素和3种荒漠植物气孔导度、 叶片水势和蒸腾速率日均值变化如表1。表层(0~30 cm)土壤含水量受降雨影响显著(P<0.05), 降雨前测得平均土壤含水量为0.035 m3·m⁻3, 降雨后测得平均土壤含水量为0.058 m3·m⁻3, 深层(30~200 cm)土壤水分变化不明显。降水前后各环境因素和3种荒漠植物的气孔导度、 叶片水势、 蒸腾速率的日变化如图1, VPD、 T和PAR在上午逐渐升高, 在13:00左右达到最大值, 在之后的测量时段VPD和T一直保持在一个较高的水平。降雨前后PAR变化很小, VPD和T变化显著(P<0.05), 其中VPD变幅最大, 主要是由于降雨后蒸发增大, 空气中水汽含量增大和气温降低共同所致。
表1 降雨前后环境因子和3种灌木气孔导度、 叶水势、 蒸腾速率日均值对比
Table 1
| 降雨前 | 降雨后 | 降雨后/降雨前 | 统计学检验P | |
|---|---|---|---|---|
| 0~30 cm土壤含水量/(m3·m⁻3) | 0.035±0.001 | 0.058±0.001 | 1.657 | 0.002* |
| 30~200 cm土壤含水量/(m3·m⁻3) | 0.064±0.001 | 0.068±0.001 | 1.063 | 0.079 |
| VPD/kPa | 5.653±0.268 | 4.602±0.283 | 0.814 | 0.000* |
| T/℃ | 36.559±1.788 | 33.769±1.384 | 0.924 | 0.000* |
| PAR/(µmol·m⁻2·s⁻1) | 891.459±35.077 | 917.385±42.508 | 1.029 | 0.724 |
| 梭梭气孔导度/(mol·m⁻2·s⁻1) | 0.096±0.008 | 0.104±0.011 | 1.083 | 0.041* |
| 泡泡刺气孔导度/(mol·m⁻2·s⁻1) | 0.113±0.011 | 0.124±0.013 | 1.097 | 0.069 |
| 沙拐枣气孔导度/(mol·m⁻2·s⁻1) | 0.125±0.012 | 0.144±0.014 | 1.152 | 0.037* |
| 梭梭叶片水势/MPa | -3.432±0.126 | -2.869±0.118 | 0.836 | 0.019* |
| 泡泡刺叶片水势/MPa | -4.706±0.154 | -3.770±0.183 | 0.801 | 0.000* |
| 沙拐枣叶片水势/MPa | -1.956±0.127 | -1.546±0.125 | 0.790 | 0.001* |
| 梭梭蒸腾速率/(mmol·m⁻2·s⁻1) | 4.572±0.248 | 4.056±0.239 | 0.887 | 0.010* |
| 泡泡刺蒸腾速率/(mmol·m⁻2·s⁻1) | 6.022±0.273 | 4.433±0.263 | 0.736 | 0.000* |
| 沙拐枣蒸腾速率/(mmol·m⁻2·s⁻1) | 6.375±0.257 | 6.099±0.243 | 0.957 | 0.101 |
*表示显著相关(P<0.05)
图1
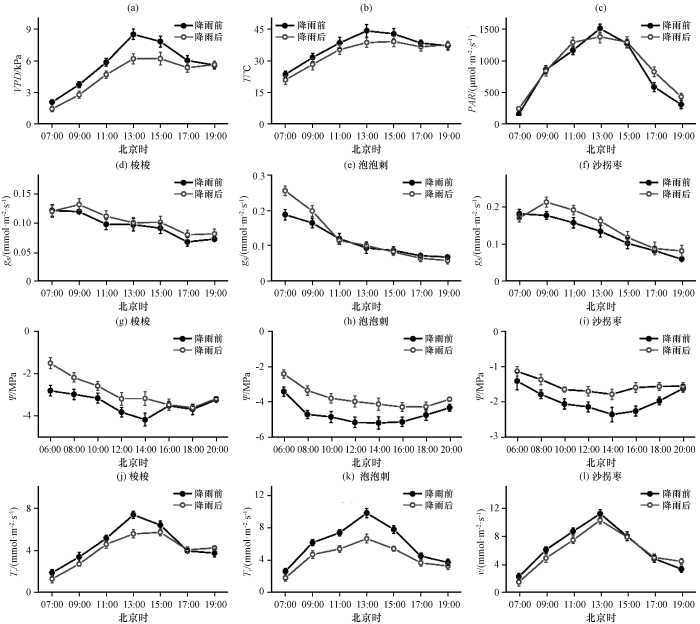
图1
降雨前后饱和水汽压差VPD (a)、 气温T (b)、 光合有效辐射PAR (c)和3种灌木气孔导度gs (d~f)、 叶水势Ψ (g~i)及蒸腾速率Tr (j~l)的平均日变化
Fig.1
Mean diurnal variations of Vapor Pressure Deficit (VPD) (a)、 air temperature (T) (b) and Photosynthetically Active Radiation (PAR) (c) and stomatal conductance (gs) (d~f)、 leaf water potential (Ψ)
气孔导度日变化表现为递减趋势, 没有出现正午气孔关闭现象, 3种植物的气孔导度均为上午最高, 之后逐渐降低, 梭梭的气孔导度在整体下降过程中还有周期性波动, 降雨后植物气孔导度升高, 这与苏培玺等对不同土壤水分处理条件下梭梭和沙拐枣响应的研究结果一致(苏培玺和严巧娣, 2006; 苏培玺, 2019)。总体而言, 梭梭的日均气孔导度最小, 泡泡刺的气孔导度最大, 沙拐枣次之。
植物叶水势在日出前最大, 在13:00左右降至最低, 之后又缓慢回升。3种荒漠植物叶片水势的大小顺序为沙拐枣>梭梭>泡泡刺, 凌晨平均水势分别为-1.274, -2.159和-2.921 MPa, 午后平均水势分别为-2.093, -3.844和-4.721 MPa, 分别降低64.3%, 78.0%和61.6%, 梭梭的叶片水势日变化最大。降雨后荒漠水分条件得到改善, 3种荒漠植物叶片水势升高(P<0.05)。凌晨水势与根区的土壤水势最接近, 分层分析根区不同深度的土壤水分与植物叶片水分状况的相关性, 结果表明梭梭的凌晨叶水势与深度为160~180 cm的土壤水分含量最相关(R2=0.919), 泡泡刺凌晨叶水势与受降雨影响的浅层土壤水分(20~40 cm)更相关(R2=0.972), 沙拐枣根区140~160 cm的土壤水分含量能比较好地反映植物凌晨叶水势情况(R2=0.808)。
3种荒漠植物蒸腾速率日变化也表现为单峰型, 与外界环境因素日变化趋势一致, 在13:00左右达到最大值, 梭梭的蒸腾速率最小, 沙拐枣的蒸腾速率最大, 泡泡刺次之, 降雨后蒸腾速率有所减少, 这与Ji et al(2016)对荒漠植物单株蒸腾速率的研究结果一致。降雨对荒漠C3植物泡泡刺叶水势增加和蒸腾速率减少的作用要大于C4植物梭梭和沙拐枣。
3.2 气孔导度对主要环境因子的响应
气孔导度的日变化主要受VPD、 T和PAR等环境因素影响, 采用2020年生长季实际观测数据对Jarvis模型的各变量分别率定, 降雨前后气孔导度对各环境驱动响应的拟合结果如表2, 对VPD的响应为反比函数(郑海雷和黄子琛, 1992), 对T的响应为二次函数, 对PAR的响应为直角双曲线函数(于贵瑞等, 2010)。3种荒漠植物的气孔导度对环境驱动因子的响应顺序均为: VPD、 T和PAR, 气孔导度主要受VPD的影响, 随着VPD的升高而逐渐降低(图2), 其对湿度的响应可以最大限度地提高植物的水分利用效率。通常气孔导度对VPD的敏感度随植物干旱或辐照度升高而增加, 荒漠C3植物泡泡刺的气孔导度随VPD增大迅速下降, 受干旱胁迫程度最高; 荒漠C4植物梭梭和沙拐枣的气孔导度对VPD的敏感度相对较低。温度为第二重要影响因子, 大气的湿度和温度直接影响环境蒸发力, 因此与气孔导度有较强的关系。降雨前后3种荒漠植物气孔导度随VPD增加而减小的响应趋势基本一致, 但降雨后相同VPD条件下对应的梭梭和沙拐枣气孔导度显著减小。气温升高, 气孔导度减小, 但由于降水稀少和湍流气温波动的影响, 降雨前后气孔导度对气温变化的响应表现出较大的不确定性。3种荒漠植物气孔导度与PAR的相关性很低, 故下文在进行气孔导度对综合环境驱动的响应模拟时没有考虑PAR。
表2 各响应函数表达式及决定系数
Table 2
| 植物 种类 | 响应函数 | 降雨前 | 降雨后 | ||||
|---|---|---|---|---|---|---|---|
| 参数 | R2 | n | 参数 | R2 | n | ||
| 梭梭 | a1=4.860, a2=6.299 | 0.393* | 64 | a1=2.345, a2=2.599 | 0.480* | 68 | |
| b1=1.502, b2=-0.049, b3=0.001 | 0.339* | 64 | b1=0.462, b2=0.013, b3=-0.001 | 0.365* | 68 | ||
| c1=0.411, c2=0.410, c3=-316.650 | 0.084 | 64 | c1=0.358, c2=0.362, c3=-577.869 | 0.020 | 68 | ||
| 泡泡刺 | a1=1.315, a2=-0.247 | 0.630* | 64 | a1=1.352, a2=0.891 | 0.770* | 68 | |
| b1=3.960, b2=-0.180, b3=0.002 | 0.684* | 64 | b1=1.199, b2=-0.027, b3=-0.001 | 0.766* | 68 | ||
| c1=0.243, c2=0.239, c3=-141.822 | 0.092* | 64 | c1=0.627, c2=0.173, c3=245.925 | 0.214* | 68 | ||
| 沙拐枣 | a1=2.768, a2=2.516 | 0.529* | 64 | a1=2.327, a2=2.770 | 0.543* | 68 | |
| b1=1.922, b2=-0.073, b3=0.001 | 0.449* | 64 | b1=-0.532, b2=0.088, b3=-0.002 | 0.476* | 68 | ||
| c1=0.141, c2=0.388, c3=434.012 | 0.012 | 64 | c1=0.384, c2=0.377, c3=-315.311 | 0.080 | 68 | ||
*表示显著相关(P<0.05)
图2
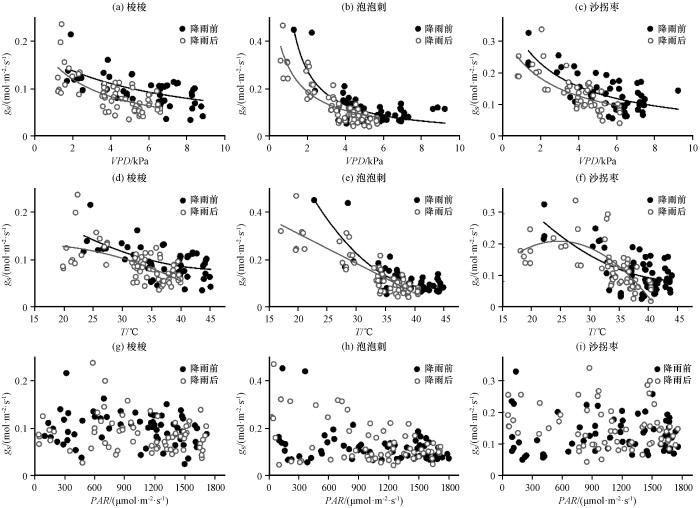
图2
降雨前后3种灌木气孔导度(gs)与饱和水汽压差(VPD) (a~c)、 气温(T) (d~f)和光合有效辐射(PAR) (g~i)的关系
Fig.2
Relationship between stomatal conductance (gs) and Vapor Pressure Deficit (VPD) (a~c)、 air temperature (T) (d~f) and Photosynthetically Active Radiation (PAR) (g~i) of the three desert shrubs prior to and following rainfall
3.3 气孔导度与叶片水势的关系
气孔导度对叶片水势的响应为线性函数(于贵瑞等, 2010), 降雨前后的拟合结果如表3, 相关性均达到了显著水平。3种荒漠植物的气孔导度随着叶水势的降低逐渐减小(图3), 随着蒸腾作用的持续进行, 叶片通过降低水势增大水势梯度以获取根区水分补给。降雨后叶片水势升高, 气孔导度随叶水势增加的斜率减小, 其中泡泡刺的变化程度最大, 对降雨的响应最为敏感。
表3 气孔导度响应叶水势的函数表达式及决定系数
Table 3
| 植物种类 | 响应函数 | 降雨前 | 降雨后 | ||||
|---|---|---|---|---|---|---|---|
| 参数 | R2 | n | 参数 | R2 | n | ||
| 梭梭 | 0.332* | 42 | 0.385* | 68 | |||
| 泡泡刺 | 0.627* | 42 | 0.548* | 68 | |||
| 沙拐枣 | 0.472* | 42 | 0.353* | 68 | |||
*表示显著相关(P<0.05)
图3
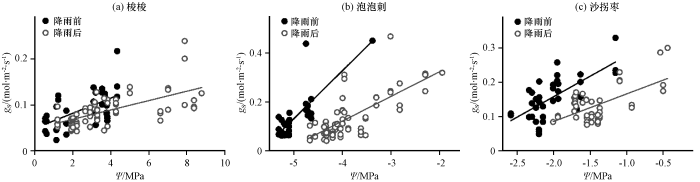
图3
降雨前后3种灌木气孔导度(gs)与叶水势(Ψ)的关系
Fig.3
Relationship between stomatal conductance (gs) and leaf water potential (Ψ) of the three desert shrubs prior to and following rainfall
3.4 不同荒漠植物气孔导度对环境因子和叶片水势的综合响应
上述结果表明: 降雨前后3种植物气孔导度对各变量的响应表现出一定的差异, 且在大多数据集中, 较湿润和较干旱的时期都存在, 而参数拟合算法倾向于在高气孔导度值下最小化较大的绝对误差, 从而将误差集中在较干旱时段的数据, 这可能会导致模拟不太准确(Anderegg et al, 2017), 故有必要将降雨前后的数据分开拟合。在两个环境变量的基础上增加了对植物水分关系的响应, 3种荒漠植物气孔导度的综合驱动因子包括VPD、 T和Ψ三个变量, 结果显示降雨前后的模拟值和实测值的相关性均达到了显著水平(表4, 图4)。3种植物中泡泡刺的拟合效果最好, 对外界环境因子变化最敏感, 梭梭和沙拐枣的拟合效果次之。3种荒漠植物的光合途径不同, 泡泡刺是C3植物, 沙拐枣和梭梭是C4植物, 在荒漠高温环境下C4植物更有优势, 水分利用效率更高, 表现为两种C4植物沙拐枣和梭梭的气孔导度较低, 相比之下, C3植物泡泡刺的气孔导度更高, 对外界水分环境变化更敏感, 与各环境因子的相关性更高, 气孔导度模型拟合效果更好。
表4 气孔导度模型表达式及决定系数
Table 4
| 植物种类 | 模型表达式 | 外界环境 | 2020年 | 2019年 | ||
|---|---|---|---|---|---|---|
| R2 | n | R2 | n | |||
| 梭梭 | 降雨前 | 0.625* | 63 | 0.479* | 90 | |
| 降雨后 | 0.722* | 65 | 0.646* | 72 | ||
| 泡泡刺 | 降雨前 | 0.747* | 63 | 0.827* | 90 | |
| 降雨后 | 0.858* | 65 | 0.682* | 72 | ||
| 沙拐枣 | 降雨前 | 0.689* | 63 | 0.669* | 90 | |
| 降雨后 | 0.674* | 65 | 0.651* | 72 | ||
*表示显著相关(P<0.05), 2020年实测数据对模型率定, 2019年实测数据对模型验证
图4
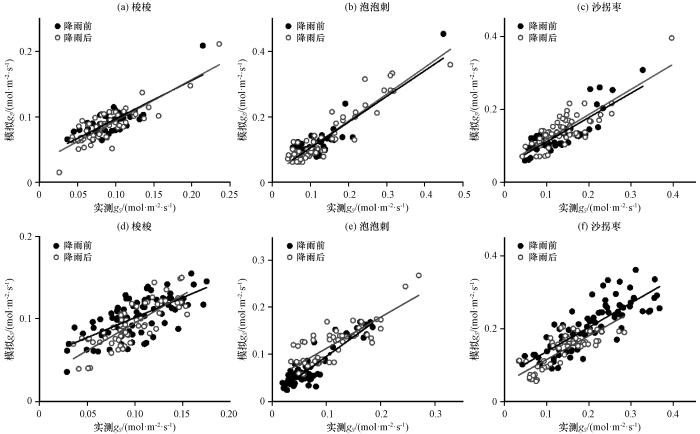
图4
2020年(a~c)和2019年(d~f)3种灌木气孔导度模拟值与实测值比较
Fig.4
Comparison between simulated and measured stomatal conductance values of the three shrubs in 2020 (a~c) and 2019 (d~f)
3.5 气孔导度和VPD对蒸腾的调控作用
3种灌木叶片蒸腾速率与气孔导度和VPD的关系如图5和表5, 受饱和水汽压差的制约, 上午蒸腾速率和气孔导度的相关性相对较弱, 随气孔导度的增大先升高后降低, 梭梭、 泡泡刺和沙拐枣的蒸腾速率对气孔导度响应的拐点分别为0.171 mol·m-2·s-1、 0.193 mol·m-2·s-1和0.257 mol·m-2·s-1, 下午虽然气孔导度较小, 但饱和水汽压差较大, 受饱和水汽压差的影响, 反而提高了蒸腾速率随气孔导度增加而增大的变化率。蒸腾速率随VPD的升高而增大, VPD在13:00达到峰值之前和之后对蒸腾速率的驱动响应不同, 上午泡泡刺和沙拐枣的蒸腾速率与VPD二次曲线的二项式系数为负数, 对蒸腾速率存在制约作用, 曲线的拐点泡泡刺为7.625 kPa, 沙拐枣为7.667 kPa, 而梭梭没有表现出这种现象, 这表明根系相对较浅的泡泡刺和沙拐枣受到土壤水分的制约, 而梭梭较深的根系受地下水影响, 根系区土壤水分供给相对充足; 下午蒸腾速率与VPD二次曲线的二项式系数为正数, 蒸腾速率随VPD的减小而下降。蒸腾速率对VPD的响应关系随时间变化, 这和植物根-叶水势梯度增大有关 (Zeppel et al, 2004)。3种灌木上午的气孔导度较高但VPD较低, 蒸腾速率主要受VPD的影响, 下午气孔导度降低, 蒸腾速率同时受到气孔导度的制约。大气水势和叶水势的水势差近似于与叶片接触的空气的饱和水汽压差, 其和蒸腾速率以及气孔导度之间的关系可以类比于欧姆定律, 3种荒漠植物Tr和VPD·gs的关系(图6), 均达到了很高的相关性, 说明气孔导度和饱和水汽压差的乘积基本可以解释蒸腾速率的变化。
图5
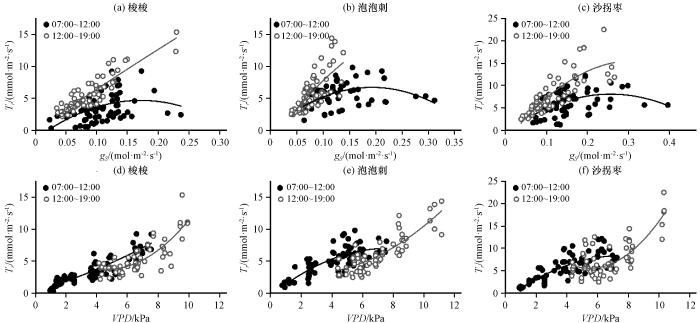
图5
蒸腾速率(Tr)与气孔导度(gs) (a~c)和饱和水汽压差(VPD) (d~f)的关系
Fig.5
Relationship between transpiration rate (Tr) and stomatal conductance (gs) (a~c) and Vapor Pressure Deficit (VPD) (d~f)
表5 蒸腾速率(Tr)与气孔导度(gs)和饱和水汽压差(VPD)的函数表达式及决定系数
Table 5
| 植物种类 | 响应函数 | 时间 | 参数 | R2 | n |
|---|---|---|---|---|---|
| 梭梭 | 07:00 -12:00 | 0.221* | 77 | ||
| 12:00 -19:00 | 0.807* | 69 | |||
| 07:00 -12:00 | 0.817* | 77 | |||
| 12:00 -19:00 | 0.662* | 69 | |||
| 泡泡刺 | 07:00 -12:00 | 0.212* | 77 | ||
| 12:00 -19:00 | 0.516* | 69 | |||
| 07:00 -12:00 | 0.631* | 77 | |||
| 12:00 -19:00 | 0.724* | 69 | |||
| 沙拐枣 | 07:00 -12:00 | 0.231* | 77 | ||
| 12:00 -19:00 | 0.680* | 69 | |||
| 07:00 -12:00 | 0.654* | 77 | |||
| 12:00 -19:00 | 0.637* | 69 |
*表示显著相关(P<0.05)
图6
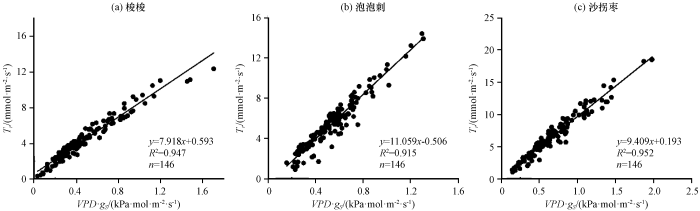
图6
蒸腾速率(Tr)与饱和水汽压差(VPD)和气孔导度(gs)乘积比较
Fig.6
Comparison of transpiration rate (Tr) with product of Vapor Pressure Deficit (VPD) and stomatal conductance (gs)
4 讨论
4.1 气孔导度-VPD-蒸腾速率关系
事实上, 蒸腾速率、 气孔导度和环境蒸发需求(VPD)之间存在复杂的相互作用和反馈机制。一方面, 叶片气孔导度会对植物水分状况变化做出响应, 而叶片水势变化则取决于蒸腾速率的大小, 随着叶水势越来越低, 气孔导度关闭的趋势进一步增加, 表现为对蒸腾作用的反馈响应(Dewar, 1995; Monteith, 1995)。另一方面, VPD增大会引起蒸腾速率的增加, 为了避免因VPD的增加而引起的蒸腾速率过高, 气孔导度会减小以避免植物水势的相应下降(Oren et al, 2010), 气孔导度减小是对空气湿度降低、 VPD增加的前馈反应, 气孔导度在水分亏缺造成不利影响前降低是植物对干旱适应的一种机制(龚吉蕊等, 2005)。
4.2 叶片水势对气孔导度的影响
目前关于水分因素中叶水势对气孔导度影响的研究有不同看法, 一些研究认为叶片水势是气孔导度的决定性影响因素, 即认为干旱条件下, 植物气孔导度的下降主要是由于叶片水势的降低(Ludlow, 1976), 而一些研究表明没有观察到气孔导度对叶水势的明显反应, 认为可能由于植物对叶片失水的敏感控制而没有达到足够低的水势导致气孔导度降低(Whitehead et al, 1981)。这与植物种类和外界环境有关, 事实上, 气孔对叶水势的响应可能是植物干旱适应策略的核心要素之一(Klein, 2014; Meinzer et al, 2016)。降雨前3种荒漠植物的气孔导度与叶水势线性关系的斜率更高, 受叶水势影响更明显, 表明在水分胁迫发展过程中, 植物对叶水势变化的气孔调节逐渐增加。叶水势反映了植物的水分亏缺情况, 影响荒漠植物气孔的张开程度, 在气孔导度估算模型中加入叶水势这一变量既提高了模型的预测能力, 也大大降低了目前经验模型在干旱条件下模拟的偏差(Anderegg et al, 2017)。
4.3 荒漠植物对土壤水分的利用
荒漠C4植物梭梭表现出低气孔导度、 低蒸腾、 低叶片水势特点, 具有忍耐脱水的能力(龚吉蕊等, 2005), 水分主要来源于深层土壤水或地下水。荒漠C3植物泡泡刺的气孔导度最高, 蒸腾速率也比较高, 叶片水势最低, 由于水分耗散速度较快, 其水分利用策略为以低水势尽可能获取水分从而保持相对高的气孔开度和蒸腾速率以应对荒漠的高温环境, 主要依赖于受季节性降水影响的浅层土壤水。荒漠C4植物沙拐枣的气孔导度较高, 而且蒸腾速率和叶片水势最高, 叶水势下降后能够较快回升, 通过利用深层土壤水保持高水势, 与大气之间形成的较高水势差造成较大的蒸腾速率以应对干旱环境。这与周海等(2017)对荒漠植物的同位素水文学研究结果和Ji et al (2016)水量平衡研究结果一致, 在类似环境条件下, 闫珂等(2020)对准噶尔盆地南缘的梭梭的研究认为在有降雨补给时, 梭梭主要利用浅层土壤水, 没有降雨补给时, 梭梭主要利用深层土壤水和地下水, 褚建民(2007)对甘肃民勤荒漠植物对水源利用策略的研究也认为梭梭和沙拐枣对土壤水的利用深度较泡泡刺深。3种灌木不同的水分利用策略是其在长期适应荒漠干旱环境的过程中形成的(徐贵青和李彦, 2009), 这会影响植物的气孔行为以及气孔导度对环境因子的响应关系。植物水分利用策略的不同也使得气孔导度模拟时土壤水深度的选取更为复杂, 因此模型中水分因子更适合采用叶水势而不是土壤水。
4.4 边界层导度对蒸腾速率的影响
叶片蒸腾对气孔导度的敏感性还与风速和边界层阻力有关, 随着空气运动增加、 边界层阻力的破坏, 蒸腾作用能更好地对气孔导度变化作出响应, 因此考虑到在叶片表面的层流副层, 即由于分子粘滞力形成的边界层阻力的影响, 在叶片气孔导度gs基础上增加叶片边界层导度(gbs, 单位: mol·m-2·s-1), 串联作为叶片表面导度(gts, 单位: mol·m-2·s-1), 模拟结果显示相关系数和斜率变化并没有预期明显(结果未展示), 说明3种荒漠植物叶片蒸腾速率受边界层导度影响较小, 气孔导度的调控占主导地位。
5 结论
通过对河西走廊中部荒漠-绿洲过渡带3种典型固沙植物梭梭、 泡泡刺和沙拐枣叶片水分交换过程的研究, 主要结论有:
(1) 3种荒漠植物的气孔导度主要受VPD的影响, 随着VPD的升高而降低, 其次气孔导度随T升高而降低, 随着叶水势的降低逐渐减小, 但与PAR的相关性很低。3种植物气孔导度对环境因子的响应与其水分利用策略和光合途径有关, 不同荒漠植物气孔导度对环境因子和叶片水势的综合响应模拟研究表明, C3植物泡泡刺对这些因子变化的响应比C4植物梭梭和沙拐枣更敏感, 与各因子的相关性更高。
(2) 应用Jarvis气孔导度模型并采用非线性响应函数构建了3种荒漠植物气孔导度对综合环境驱动的响应模型, 能够很好地模拟气孔导度的日内变化, 这为估算荒漠植物蒸腾过程提供了重要参考。
(3) 3种荒漠植物下午气孔导度对蒸腾的调控作用较上午高, 考虑到VPD的影响, 通过与电路类比, 结果显示气孔导度和饱和水汽压差的乘积基本可以解释蒸腾速率的变化, 在此基础上又考虑了叶片边界层导度的影响, 模拟结果没有明显变化, 说明气孔导度是控制3种荒漠植物叶片蒸腾的主要机制。
参考文献
Plant water potential improves prediction of empirical stomatal models
[J].
Stomata: Key players in the earth system, past and present
[J].
Carbon-water balance and patchy stomatal conductance
[J].
Maximum rooting depth of vegetation types at the global scale
[J].
How plants cope with water stress in the field?Photosynthesis and growth
[J].
Interpretation of an empirical model for stomatal conductance in terms of guard cell function
[J].
Non-convergent transpiration and stomatal conductance response of a dominant desert species in central Asia to climate drivers at leaf, branch and whole plant scales
[J].
Global diurnal and nocturnal parameters of stomatal conductance in woody plants and major crops
[J].
The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field
[J].
Transpiration from three dominant shrub species in a desert-oasis ecotone of arid regions of northwestern China
[J].
Stomatal control of photosynthesis and transpiration
[J].
Maximum conductances for evaporation from global vegetation types
[J].
The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviors
[J].
Ecophysiology of C4 Grasses
[J].
Mapping 'hydroscapes' along the iso-to anisohydric continuum of stomatal regulation of plant water status
[J].
A reinterpretation of stomatal responses to humidity
[J].
Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit
[J].
Drylands in the earth system
[J].
Carbon dioxide and water vapor exchange in response to drought in the atmosphere and in the soil
[J].
Predicting stomatal responses to the environment from the optimization of photosynthetic gain and hydraulic cost
[J].
Stomatal response to environmental variables in two tropical forest species during the dry season in Nigeria
[J].
Root distribution of three dominant desert shrubs and their water uptake dynamics
[J].
Leaf temperature fluctuations of typical psammophytic plants and their application to stomatal conductance estimation
[J].
Seasonal responses of xylem sap velocity to VPD and solar radiation during drought in a stand of native trees in temperate Australia
[J].
Water-use response to climate factors at whole tree and branch scale for a dominant desert species in central Asia: Haloxylon ammodendron
[J].
干旱区植物的水分选择性利用研究
[D].
蒸散发模型结合微气象数据模拟陆面蒸散发研究进展
[J].
黑河流域几个主要植物种光合特征的比较研究
[J].
多年冻土区土壤蒸散发对气候变化的敏感性分析
[J].
植物应对干旱胁迫的气孔调节
[J].
荒漠河岸林胡杨光合参数变化特征及影响因子研究
[J].
C4荒漠植物梭梭和沙拐枣在不同水分条件下的光合作用特征
[J].
荒漠植物梭梭和沙拐枣光合作用、 蒸腾作用及水分利用效率特征
[J].
古尔班通古特沙漠不同生活型植物对小雨量降雨的响应
[J].
黑河中下游不同类型下垫面的能量收支差异及其成因研究
[J/OL].
共生条件下三种荒漠灌木的根系分布特征及其对降水的响应
[J].
3种荒漠灌木的用水策略及相关的叶片生理表现
[J].
准噶尔盆地南缘梭梭水分来源与传输规律分析
[J].
中国温带荒漠植物蒸腾过程模拟的若干问题分析
[J].
古尔班通古特沙漠西部地下水位和水质变化对植被的影响
[J].
春小麦单叶气孔行为及蒸腾作用的模拟
[J].
两种荒漠生境条件下泡泡刺水分来源及其对降水的响应
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}